農業における誘引の本当の意味を知っていますか?単に縛るだけではなく、光合成や病気予防、収穫量に直結する重要な作業です。この記事では、基礎知識から紐の結び方、植物ホルモンとの関係まで徹底解説します。あなたの畑の野菜、正しく誘引できていますか?


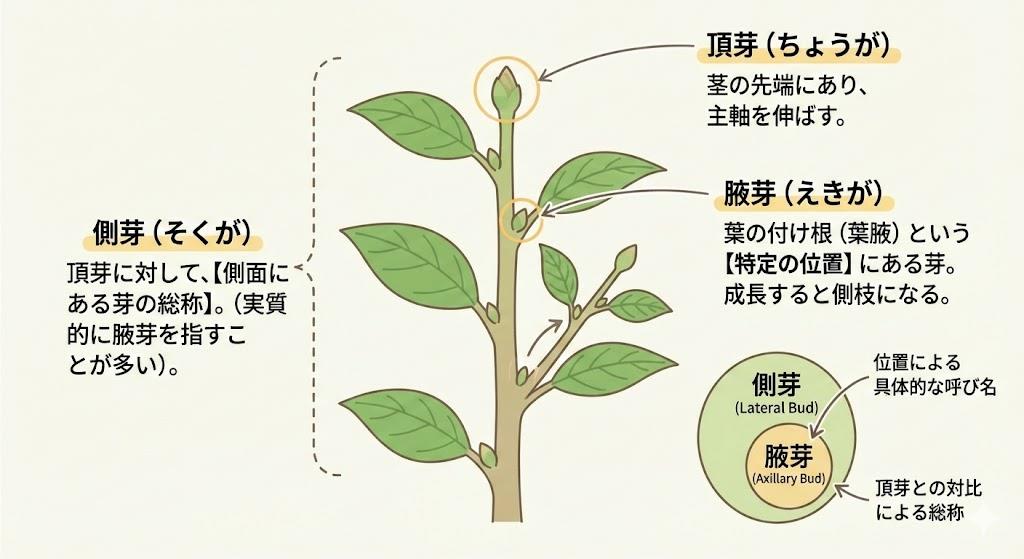
植物が上へ向かって優先的に成長する現象、それが「頂芽優勢」です 。この現象の主役となっているのが、「オーキシン」という植物ホルモンです。オーキシンは、主に茎の先端にある「頂芽」で活発に合成されます 。合成されたオーキシンは、重力に従って植物体内を上から下へと移動していく性質があります。このオーキシンの流れが、側面にある「わき芽(側芽)」の成長を抑制する働きを持っています 。
具体的には、頂芽で高濃度に保たれたオーキシンが、わき芽の細胞分裂や伸長を直接的に、あるいは間接的に抑え込むことで、頂芽自身の成長を最優先させています 。これにより、植物は競争相手よりも早く太陽光を確保するために、まず高さを稼ぐという生存戦略をとっていると考えられています。もし頂芽が動物に食べられたり、剪定されたりして失われると、オーキシンの供給が止まります。その結果、わき芽を抑制していた力がなくなり、わき芽が一斉に成長を開始します。これが、剪定後に枝数が増える基本的な仕組みです 。
オーキシンの働きは、単にわき芽を抑制するだけではありません。以下の表のように、植物の成長において多様な役割を担っています。
| オーキシンの主な働き | 農業における応用例 |
|---|---|
| 細胞の伸長促進 | 果実の肥大促進剤としての利用 |
| 発根の促進 | 挿し木や挿し穂の発根剤として利用 |
| 落葉・落果の抑制 | 収穫前の果実の落下防止 |
このように、頂芽優勢の仕組みを理解することは、植物の基本的な生育メカニズムを知る上で非常に重要であり、日々の農作業における多くの技術的判断の根拠となります 。
以下のリンクは、大学の研究室によるQ&Aで、頂芽優勢におけるオーキシンとサイトカイニンの関係性について、より専門的な見地から解説されています。
頂芽優勢はオーキシンだけで決まるわけではありません。もう一つの重要な植物ホルモン、「サイトカイニン」が深く関わっています 。サイトカイニンは、主に根の先端で合成され、導管を通って地上部へと供給されるホルモンです。その主な役割は、細胞分裂を促進し、わき芽の成長を促すことです 。つまり、オーキシンがわき芽を「抑制する」のに対し、サイトカイニンは「促進する」という、正反対の働きを持っています。
頂芽優勢の仕組みは、この二つのホルモンの「濃度バランス」によって絶妙に制御されています。頂芽に近い部分では、頂芽から供給される高濃度のオーキシンが、サイトカイニンの合成や働きを抑制します 。そのため、サイトカイニンが存在していても、わき芽は成長することができません。一方で、根に近い株元のわき芽は、頂芽から遠いためにオーキシンの濃度が低く、根から供給されるサイトカイニンの影響を強く受けるため、成長しやすくなります。これが、多くの植物で下の方の枝がよく茂る理由の一つです。
このオーキシンとサイトカイニンの濃度比(オーキシン/サイトカイニン比)が、植物の各部位が「芽」になるか「根」になるかを決定づける重要な要素であることも知られています。
この原理は、植物の組織培養技術において、特定の器官を再生させるために広く応用されています。農業現場においても、例えば窒素肥料を適切に施用することは、根の活動を活発にし、サイトカイニンの合成を促すことにつながります。これが間接的にわき芽の発生や生育に影響を与える可能性があることは、あまり知られていない事実かもしれません。最新の研究では、サイトカイニンの新たな活性化経路も発見されており、この遺伝子を人為的に調節することで、イネなどの作物の収量を向上させる未来が期待されています 。
農業生産において、頂芽優勢の原理は収量や品質を向上させるための重要な技術として広く応用されています 。代表的な作業が「摘心(てきしん)」や「剪定(せんてい)」です。
摘心とは、主枝の先端にある頂芽を摘み取る作業のことです 。頂芽を摘むことでオーキシンの供給源がなくなり、頂芽優勢が打破されます。これにより、それまで抑制されていたわき芽が一斉に成長を始め、枝数が増加します。ナスやキュウリなどの果菜類でこの技術を用いることで、花(=果実)がつく側枝を増やすことができ、結果的に収穫量を大幅に増やすことが可能になります。
剪定も同様の原理に基づいています 。不要な枝や古い枝を切り詰めることで、
また、トマトの栽培で行われる「わき芽かき」は、逆の発想に基づいています。トマトは生育旺盛で次々とわき芽を出しますが、これを放置すると栄養が分散してしまい、主軸の果実が大きく育ちません。そこで、あえてわき芽を小さいうちに取り除くことで、オーキシンの働きを維持し、栄養を特定の花や果実に集中させて大玉で高品質なトマトを収穫するのです。このように、頂芽優勢の仕組みを科学的に理解することは、作物の種類や栽培目的に応じて、摘心、剪定、わき芽かきといった管理作業をより効果的に行うための羅針盤となります。
頂芽優勢の制御は、オーキシンとサイトカイニンだけで完結する単純なものではありません。実は、他の植物ホルモンも複雑に絡み合い、相互に作用することで、より精密な成長制御ネットワークを形成しています。これは一般的にはあまり知られていない、専門的な視点です。
ジベレリン: ジベレリンは、主に植物の伸長成長を促進するホルモンとして知られています 。種なしブドウの生産に使われることで有名です 。頂芽優勢との関連では、ジベレリンはオーキシンと協力して茎の伸長を促す一方で、わき芽の休眠打破にも関与することが示唆されています。オーキシンの濃度が低下した際に、ジベレリンがわき芽の成長開始の引き金の一つとなる可能性があるのです。
エチレン: エチレンは、果実の成熟や老化(葉の黄化など)を促進するガス状のホルモンです。ストレス条件下で多く生成されることでも知られています。頂芽優勢の文脈では、高濃度のオーキシンがエチレンの合成を誘導し、そのエチレンがわき芽の成長を抑制するという説があります。つまり、オーキシンがエチレンを介して間接的にわき芽を抑えている可能性があるのです。
ストリゴラクトン: 比較的新しく発見された植物ホルモンで、頂芽優勢の制御において非常に重要な役割を担っていることが明らかになってきました 。ストリゴラクトンは、根から地上部へ、そして茎の中でも輸送され、オーキシンと協調してわき芽の成長を強力に抑制します。頂芽から供給されるオーキシンが、わき芽でのストリゴラクトンの感受性を高める、あるいはストリゴラクトンそのものの作用を強めることで、抑制効果が発揮されると考えられています。頂芽を切除するとオーキシンの供給が止まり、ストリゴラクトンの抑制効果が解除され、サイトカイニンの働きでわき芽が成長するという、より精緻なモデルが提唱されています。さらに、ストリゴラクトンは土壌中の微生物との共生を促すシグナルとしても機能しており、植物の栄養状態とも密接に関連しています 。
このように、頂芽優勢という一つの現象をとっても、複数のホルモンがオーケストラのように協調し、あるいは拮抗しながら、植物の形づくりを巧みにコントロールしています。
アースガーデン 除草剤 アースカマイラズ 草消滅 [4.5L] ガーデニング ジョウロヘッド(シャワー タイプ) 家庭用 園芸 雑草 除去 除草剤 (アース製薬)